
曾琼戎, 刘泽峰, 王文娟, 黄雪莲, 黎凤炎, 夏雨艳, 张 国
肝细胞癌(hepatocellular carcinoma,HCC)的发病率和病死率在国内外均呈上升趋势[1-2],其中超过半数的病例发生在我国。HCC的治疗受到较多局限性,除了患者确诊较晚外,还与HCC的异质性、化疗耐受等特点相关。索拉非尼(sorafenib)作为一种酪氨酸激酶抑制剂(tyrosine kinase inhibitors,TKI),长期以来一直是唯一被批准用于晚期HCC治疗的药物,但大多数患者显示出原发性或获得性耐药,使其疗效受限[3]。近年,也有其他多激酶抑制剂(乐伐替尼、卡博替尼、瑞戈非尼)、人单克隆抗体(雷莫芦单抗)和免疫检查点抑制剂(纳武单抗、派姆单抗)被批准用于全身治疗[4-6]。但尽管如此,患者的中位生存期并未显著增加[7]。因此,迫切需要了解HCC索拉非尼耐药性的分子机制,以提高当前药物的疗效,并开发更有效的药物。为此,本课题组在体外成功构建了两种耐索拉非尼肝癌细胞株,并对索拉非尼的耐药机制进行了探索,为HCC治疗提供理论模型和基础。
1.1主要实验试剂 索拉非尼(MCE,HY-10201),高糖DMEM培养基(Gibco,C11995500BT),澳洲胎牛血清(fetal bovine serum,FBS;
Gibco,10099-141),青链霉素(penicillin-streptomycin,PS;
索莱宝,PI400-100),CCK8试剂盒(MCE,HY-K0301),Annexin V/PI凋亡检测试剂盒(赛默飞,88-8005-72),TRIzol(赛默飞,15596-026),逆转录试剂盒(莫纳,MR05101),SYBR Green qPCRMix(莫纳,MQ10301S)。兔单克隆一抗:caspase-3、自噬受体蛋白P62、LC3、β-actin(Abcam公司)。山羊抗兔二抗(Affinity公司)。PCR所用引物均由上海生工设计。
1.2野生型HCC细胞株HepG2、Huh7的培养 野生型人肝癌细胞株HepG2、Huh7,购自中国科学院上海细胞所,以含10% FBS+1% PS的DMEM培养基进行细胞培养,培养箱条件设置为37 ℃、5% CO2。
1.3耐索拉非尼人肝癌细胞株的建立 采用浓度递增法构建耐药株,具体流程如下:稳定培养野生型人肝癌细胞株HepG2、Huh7至P3代,密度达30%时进行传代,贴壁后加入含3 μM索拉非尼的完全培养基(含15% FBS)。培养3 d后更换为不含索拉非尼的完全培养基,待细胞长满后,再次以30%进行传代。细胞贴壁后加入4 μM索拉非尼,培养3 d后更换成不含索拉非尼的完全培养基,待细胞长满再次以30%进行传代。细胞贴壁后加入5 μM索拉非尼,培养3 d后更换成不含索拉非尼的完全培养基。待细胞长满后再以30%进行传代。以此类推,直至加入含10 μM索拉非尼的完全培养基经3 d培养细胞可长满。之后以含5 μM的索拉非尼的完全培养基维持培养1个月以上,所得细胞株即耐索拉非尼人肝癌细胞株(HepG2-SR和Huh7-SR),以含15% FBS+1% PS的DMEM培养基进行培养,并用于后续实验。
1.4CCK8实验方法 取对数生长期细胞株HepG2、Huh7、HepG2-SR和Huh7-SR,以5 000 cells/孔接种至96孔板,培养过夜,更换含梯度浓度索拉非尼(2.5、5.0、10.0、20.0 μM)的完全培养基,另设等体积不加药物处理的阴性对照组和空白对照组,每组3个复孔。培养24 h后,小心吸弃旧培养基,然后向每个孔加入90 μl的10 % FBS DMEM和10 μl的CCK8试剂,37 ℃培养箱继续孵育3 h。应用酶标仪检测每个孔450 nm处的吸光度值(OD值)。细胞存活率=(实验组OD值-空白组OD值)/(对照组OD值-空白组OD值)×100%。
1.5细胞形态学观察 将细胞接种于6 cm培养皿中,密度为2×105cells/孔,在附有电荷耦合原件图像传感器的显微镜下观察细胞轮廓和形态,并拍照记录。
1.6流式细胞术检测细胞凋亡 按照Annexin V/PI凋亡检测试剂盒说明书进行细胞凋亡检测。取生长良好的细胞株HepG2、Huh7、HepG2-SR和Huh7-SR接种至6孔板中,贴壁后以5 μM索拉非尼干预24 h,收获细胞并用PBS洗涤2次,采用胰酶消化细胞,室温下1 000 r/min离心5 min,吸弃上清液。每管加入2 ml PBS,重悬洗涤细胞,再次离心(1 000 r/min,5 min)吸弃上清液。用1×binding buffer缓冲液洗涤、离心1次后每管加入1×binding buffer溶液400 μl、AnnexinⅤ-FITC 5 μl,室温下避光染色10~15 min。上机前5 min加入5 μl PI染色5min,再补加200 μl 1×binding buffer溶液,2~8 ℃避光孵育约2 h,应用流式细胞仪进行分析检测。
1.7Western blot实验 取生长良好的细胞株HepG2、Huh7、HepG2-SR和Huh7-SR接种至6孔板中,贴壁后以5 μM索拉非尼干预24 h,应用RIPA裂解液提取细胞总蛋白。电泳后将SurePAGE凝胶上的条带转至聚偏二氟乙烯(polyvinylidene difluoride,PVDF)膜,在室温下用5%脱脂奶粉液封闭2 h,用TBST洗膜3次,5 min/次。在4 ℃下孵育一抗过夜,TBST洗涤3次,8 min/次。室温下孵育二抗1 h,用TBST洗涤3次,10 min/次。应用增强化学发光检测系统扫描,以β-actin为内参计算目标蛋白的相对表达量。
1.8实时荧光定量RT-PCR 取生长良好的细胞株HepG2、Huh7、HepG2-SR和Huh7-SR接种至6孔板中,贴壁后以5 μM索拉非尼干预24 h。通过TRIzol法提取细胞总RNA,按照逆转录试剂盒说明书操作步骤将RNA逆转录成cDNA,并以cDNA为模板进行实时荧光定量RT-PCR,反应体系为:MonAmp GreenTMqPCR Mix 10 μl,上下引物各0.4 μl,dH2O 7.2 μl,cDNA 2 μl。反应程序:95 ℃预变性30 s;
90 ℃ 10 s,60 ℃ 30 s扩增40个循环。所用引物序列见表1。以GAPDH为内参,应用2-△△Ct法计算目标因子的相对表达量。

表1 引物序列

2.1索拉非尼耐药细胞株的构建结果 CCK8实验结果显示,索拉非尼对HepG2-SR、HepG2、Huh7-SR、Huh7细胞株均有抑制作用,且随着干预药物浓度升高,细胞存活率逐渐降低。经索拉非尼干预24 h后计算4组细胞株的半数抑制浓度(half inhibitory concentration,IC50),结果显示,HepG2-SR细胞的IC50显著高于HepG2细胞[(15.74±0.38)μM vs (7.27±0.44)μM;
t=5.450,P<0.001];
Huh7-SR细胞的IC50显著高于Huh7细胞[(11.73±0.27)μM vs (4.92±0.31)μM;
t=4.807,P<0.001]。提示索拉非尼耐药细胞株构建成功。见图1。

ⓐ不同浓度索拉非尼干预下HepG2-SR、HepG2细胞的存活率对比;
ⓑ不同浓度索拉非尼干预下Huh7-SR、Huh7细胞的存活率对比;
ⓒHepG2-SR与HepG2细胞株对索拉非尼的IC50比较;
ⓓHuh7-SR与Huh7细胞株对索拉非尼的IC50比较;
*P<0.05图1 CCK8实验结果图
2.2索拉非尼耐药细胞与野生型细胞的形态比较
在倒置显微镜下可见索拉非尼耐药细胞株的形态与野生型细胞株比较发生了明显变化,HepG2呈上皮样的片状形态,而HepG2-SR形态上呈明显的梭形;
Huh7呈上皮样,而Huh7-SR细胞边缘也呈现出不规则的扩张。见图2。

图2 四组细胞光学显微镜下所见(×200)
2.3索拉非尼耐药细胞与野生型细胞的迁移能力比较 细胞划痕实验结果显示,与0 h时间点为对照,48 h后观察到索拉非尼耐药细胞的迁移愈合速率比野生株更快,提示索拉非尼耐药细胞的迁移能力更强。见图3。

图3 细胞划痕实验结果图(×100)
2.4索拉非尼耐药细胞与野生型细胞的凋亡率及caspase-3蛋白表达水平比较 流式细胞术结果显示,经5 μM索拉非尼干预24 h后,索拉非尼耐药细胞的细胞凋亡率均显著低于其野生型细胞[HepG2-SR vs HepG2:(6.07±0.12)% vs (9.53±0.18)%;
t=12.860,P=0.002。Huh7-SR vs Huh7:(8.52±0.20)% vs (14.71±0.33)%;
t=10.005,P=0.008]。见图4ⓐ。另外,Western blot实验结果显示,HepG2-SR细胞和Huh7-SR细胞的caspase-3的蛋白表达水平也分别低于HepG2细胞和Huh7细胞(t=3.980,P=0.016;
t=5.497,P=0.005)。见图4ⓑ。提示耐药细胞的抗索拉非尼凋亡能力比野生细胞更强。

ⓐ流式细胞术实验结果;
ⓑWestern blot实验结果;
*P<0.05图4 索拉非尼耐药细胞和野生型细胞的凋亡率及caspase-3蛋白表达水平比较图
2.5索拉非尼耐药细胞与野生型细胞的自噬相关因子表达水平比较 实时荧光定量RT-PCR结果显示,相比于野生型细胞HepG2、Huh7,HepG2-SR细胞和Huh7-SR细胞的P62 mRNA表达水平更低,LC3 mRNA表达水平均更高,差异有统计学意义(P<0.05)。Western blot实验结果也显示,相比于野生型细胞HepG2、Huh7,HepG2-SR细胞和Huh7-SR细胞的P62蛋白表达水平更低,LC3-Ⅰ向LC3-Ⅱ转化增加,LC3-Ⅱ/LC3-Ⅰ的相对表达量升高,差异有统计学意义(P<0.05)。提示耐药细胞比野生型细胞具有更高的自噬水平,HCC的索拉非尼耐药机制可能与自噬途径有关。见图5。
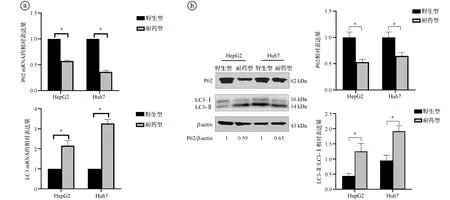
ⓐ实时荧光定量RT-PCR实验结果;
ⓑWestern blot实验结果;
*P<0.05图5 索拉非尼耐药细胞与野生型细胞的自噬相关因子表达水平比较图
3.1HCC仍是全球常见癌症相关死亡病因之一。近年来,免疫转化治疗在中晚期HCC患者中有重大突破[8]。索拉非尼作为靶向多种TKI(血管内皮生长因子受体、血小板衍生生长因子受体、丝氨酸/苏氨酸蛋白激酶家族)的抑制剂,可作用于众多靶点以抑制癌细胞增殖和肿瘤血管生成,可有效降低HCC患者的死亡率[9]。然而,临床试验表明只有约30%的患者对索拉非尼敏感,并且由于对索拉非尼耐药,HCC通常会在6个月内进展[10]。因此,建立索拉非尼耐药模型有助索于拉非尼耐药机制的研究,以开发能够增强HCC对索拉非尼敏感性的方法。
3.2本研究通过浓度递增法成功构建了HepG2-SR、Huh7-SR两种耐索拉非尼肝癌细胞系;
CCK8、流式细胞术实验结果表明,野生型和耐药细胞株均对索拉非尼有剂量依赖性,并且相较于野生型细胞株,耐药细胞株具有更强的迁移性、抗抑制性、抗凋亡等特点,提示耐药细胞株HepG2-SR、Huh7-SR构建成功。这也说明临床中长期使用索拉非尼治疗的HCC患者在治疗后期容易出现耐药性,从而影响抗癌效果。CCK8实验结果显示,HepG2-SR和Huh7-SR对索拉非尼的IC50分别为(15.74±0.38)μM和(11.73±0.27)μM,相较于李明远等[11]构建的耐药细胞模型具有更强的耐药性。
3.3在肿瘤细胞耐药性鉴定中,耐药相关蛋白鉴定也是其中之一。P-糖蛋白是一种多药转运蛋白,在肿瘤多药耐药性(multidrug resistance,MDR)中起主要作用。ATP结合盒转运蛋白G2(ATP binding cassette transporter G2,ABCG2)是ATP结合盒超家族成员之一,也是一种P-糖蛋白。有研究发现,P-糖蛋白在多种耐药肿瘤中表达水平上调[12],且ABCG2在获得性索拉非尼耐药株中呈高表达[13]。耐药相关基因在不同肿瘤中有着各自的表达特点,探寻不同肿瘤耐药基因的共同规律也许能为今后制定更为有效的恶性肿瘤MDR的分子癌症治疗提供参考。本研究Western blot实验结果显示,凋亡相关蛋白caspase-3在耐药细胞株中的剪切作用减少。在其他癌症研究中已发现通过抑制KLF5/Bcl-2/caspase-3信号通路可恢复癌细胞对化疗药物的敏感性[14],这或许能成为解决索拉非尼耐药的突破口。本研究还发现自噬标志蛋白P62及LC3的表达水平在耐药细胞株和野生型细胞株间存在差异,P62在耐药细胞株中表达下调,且LC3-Ⅱ自噬流增加,提示耐药细胞株有着更高的自噬通量。自噬与多种疾病的转归具有关联性,而P62作为一种选择性自噬受体,其活性的扰动也与许多肝病的发病机制相关[15],包括酒精性脂肪性肝炎(alcoholic steatohepatitis,ASH)、非酒精性脂肪性肝炎(non-alcoholic steatohepatitis,NASH)和HCC等。许多研究团队也正尝试将P62作为预测性生物标志物和治疗肝脏疾病的潜在靶点[16-17],但在HCC中的研究进展仍缓慢。LC3在自噬过程中发挥了重要的调控作用,LC3被半胱氨酸蛋白酶ATG4裂解形成胞质异形体LC3-Ⅰ,接着LC3-Ⅰ通过ATG7及ATG3参与泛素样反应,与磷脂酰乙醇胺(PtdEth,PE)结合,最终转化为LC3-Ⅱ,促进自噬体膜扩增[18]。因此,LC3-Ⅰ 和LC3-Ⅱ转化是判断自噬流改变的重要指标[19-20]。在本研究中,耐药株LC3-Ⅱ/LC3-Ⅰ的蛋白表达升高,提示耐药细胞株的自噬水平比野生型细胞株更高。有研究表明,自噬可以通过影响多种化疗药物的协同作用来提高对HCC细胞的致死率[21]。索拉非尼可诱导HCC保护性自噬反应的激活[22-23]。然而,索拉非尼诱导的自噬作为促生存反应并非一成不变的,也有研究在HCC中观察到相反的结果。目前,自噬和索拉非尼耐药的机制仍然有待进一步阐明,笔者将就此继续深入研究。本实验研究也存在着一些不足,实验使用的细胞株及评估自噬水平检测方法单一,且实验所用的两种肝癌野生细胞株均不是来源于我国患者,因此后续需要使用多种肝癌细胞株进行实验,同时可增加透射电镜实验以检测细胞自噬小体的生成情况。
综上所述,本研究成功构建了两种耐索拉非尼肝癌细胞株HepG2-SR、Huh7-SR,并初步探讨了HCC对索拉非尼可能的耐药机制,为HCC治疗方案的开发提供参考。
猜你喜欢索拉非尼贴壁细胞株大豆异黄酮协同索拉非尼对宫颈癌Hela细胞增殖、迁移的影响及机制新乡医学院学报(2022年10期)2022-11-07高硫煤四角切圆锅炉贴壁风倾角对水冷壁 高温腐蚀影响研究热力发电(2022年3期)2022-03-25具有一般反应函数与贴壁生长现象的随机恒化器模型的全局动力学行为数学物理学报(2021年6期)2021-12-21660MW超超临界锅炉高速贴壁风改造技术研究能源工程(2021年2期)2021-07-21索拉非尼治疗肝移植后肝细胞癌复发的单中心回顾性分析天津医科大学学报(2021年1期)2021-01-26索拉非尼治疗晚期肾癌期间引发高血压的分析中国继续医学教育(2015年3期)2016-01-06稳定敲低MYH10基因细胞株的建立医学研究杂志(2015年11期)2015-06-10Rab27A和Rab27B在4种不同人肝癌细胞株中的表达中国当代医药(2015年16期)2015-03-01稳定抑制PAK2蛋白表达的HUH—7细胞株的建立中国医药导报(2015年27期)2015-02-28体外全骨髓贴壁法培养人骨髓间充质干细胞的实验研究西南军医(2015年4期)2015-01-23版权所有:上派范文网 2010-2024 未经授权禁止复制或建立镜像[上派范文网]所有资源完全免费共享
Powered by 上派范文网 © All Rights Reserved.。沪ICP备12033476号-1